Menu
e-shop





Kvasinky jsou přirozenou součástí populace epifytních mikroorganizmů, které se běžně vyskytují na rostlinách a s nimi jsou následně zanášeny i do siláží a senáží. Největší problémy způsobují kvasinky u glycidových krmiv, jakými jsou kukuřičné siláže, cereální siláže nebo vlhké kukuřičné zrno.
Populace kvasinek a jejich metabolity se výrazně profilují především v aerobním prostředí (kyslík). Druhové složení jednotlivých populací kvasinek závisí na podmínkách během silážování. Siláž je vystavena vzdušnému kyslíku nejdříve na začátku silážního procesu a následně až po otevření siláže, v uzavřené siláži pak převažují anaerobní podmínky. Z tohoto pohledu je možno kvasinky rozdělit na druhy dominující na čerstvé píci, druhy aktivní v uzavřené siláži a konečně na druhy typické pro otevřenou siláž. Zároveň je možno kvasinky členit podle jejich schopnosti využívat rozdílné substráty, především rozpustné cukry nebo kyselinu mléčnou.
Kvasinky metabolizující cukry dominují během aerobní fáze začátku silážního procesu a během anaerobních podmínek v uzavřené jámě či vaku. Naproti tomu kvasinky využívající kyseliny vytvářejí majoritní populace na silážních stěnách, tedy za přítomnosti kyslíku. Při sklizni jsou na rostlinách tvořeny populace kvasinek z více než 90% kvasinkami metabolizujícími cukry. Vlivem silného selekčního tlaku během fermentace siláže se dominance těchto kmenů postupně snižuje, takže po otevření siláže již dominují (více než 90% zastoupení) kmeny metabolizující laktáty (Dennis 2007). Vysoké zastoupení kvasinek, které spotřebovávají kyselinu mléčnou, postupně zvyšuje pH siláže a vytváří prostředí vhodné pro růst hnilobných bakterií a plísní.
Hlavními produkty kvasinek v aerobních podmínkách jsou teplo, oxid uhličitý a kyselina octová. Zvýšená teplota a její sekundární efekty mohou ovlivnit chutnost krmiva, oxid uhličitý zvyšuje silážní ztráty sušiny. Přestože se kvasinky v anaerobním prostředí nerozmnožují, zůstávají metabolicky aktivní a produkují teplo, oxid uhličitý, etanol a další vedlejší produkty včetně kyseliny octové, aldehydů a esterů (Dennis 2007). Produkce výše zmíněného alkoholu nemusí být jednoznačně negativním jevem. Etanol pomáhá rozpouštět bílkovinu zein kukuřičného zrna a zvyšuje stravitelnost škrobu během silážování (Owens a Soderlund 2007; Hoffman 2007).
Vedle kyseliny octové a limitovaného množství etanolu produkují kvasinky aerobních podmínkách i menší množství aromatických sloučenin. Produkce těchto sloučenin je závislá na prostředí a na konkrétních druzích zastoupených kvasinek. Pokud se teplota prostředí zvyšuje, zvyšuje se obvykle i produkce těchto aromatických sloučenin. Kvasinky mohou produkovat estery (ovocné aroma), etylacetát (aroma laku na nehty), směsi kyselin a vyšších alkoholů (v lihovarnictví jsou označovány jako přiboudlina s typickým aroma ředidla), aldehydy (máselné aroma), acetaldehydy (aroma zeleného jablka) nebo další sloučeniny s typickým zápachem (Dennis 2007).
Složení substrátu ovlivňuje tvorbu vedlejších produktů také v anaerobním prostředí. Zvýšená teplota a hladina cukrů zvyšuje produkci aromatických esterů a vyšších alkoholů. Vysoká hladina cukrů může nadřadit produkci alkoholů nad ostatní metabolity. Produkce těchto aromatických sloučenin v siláži nezvyšuje pouze ztráty sušiny, ale může významně přispět k problémům s chutností krmiva (Dennis 2007).
Aerobně změněné siláže mají obvykle populace kvasinek převyšující 100 000 cfu/g hmoty. Výskyt kvasinek Hansula a Candida je obvykle spojený se zvýšeným pH, protože tyto druhy spotřebovávají kyselinou mléčnou. Naproti tomu kvasinky druhu Torulopsis primárně spotřebovávají rozpustné cukry, a proto nemají vliv na pH siláže. Většinou je však pro působení „aerobních“ kvasinek typické, že mění profil těkavých masných kyselin siláže, dochází k redukci obsahu kyseliny mléčné a ke zvýšení hladiny kyseliny octové. Nicméně vzorky odebrané z hlubších partií siláže často vykazují optimální pH i obsah kyseliny mléčné, protože je růst kvasinek v těchto místech omezen nedostatkem kyslíku (Mahanna a Chase 2003).
Jak tedy zabránit růstu a množení kvasinek v siláži? Zásadní je, co nejrazantněji omezit okysličování siláže na možné minimum. To znamená: zvýšit denní odběr siláže ze stěny, což umožní rychleji a průběžně odstraňovat rostoucí kvasinky z povrchu stěny, udržovat stěnu siláže hladkou, nenarušovat zhutnění siláže nevhodnou technikou odběru a dbát na pořádek v silážní jámě. Z dlouhodobého hlediska je nutné správně plánovat velikost skladovacích prostor tak, aby byl zajištěn dostatečně velký a pravidelný denní odběr siláže, co nejrychleji naskladnit a uzavřít jámu, sklízet ve správné zralosti a sušině, zajistit dostatečné zhutnění silážované hmoty a její utěsnění, siláž stabilizovat silážními aditivy obsahujícími kmen Lactobacillus buchneri.
V této souvislosti je nutné si připomenout, že optimální doba zrání siláže je 6–8 týdnů. Předčasné zkrmování nedostatečně vyzrálé siláže je z dietetického hlediska rizikové a může mít negativní dopad na zdravotní stav a užitkovost zvířat. Navíc jsou tyto siláže velmi aerobně nestabilní, a tím i náchylné k druhotnému zahřívání a rozvoji patogenních mikroorganismů.
Řešení nabízí inovované konzervanty firmy Pioneer, které jsou schopny kvasný proces urychlit, a tím dát možnost zkrmovat siláže výrazně dříve. Otevřít siláže již za deset dní po jejich zasilážování umožňují tzv. Rapid React™ inokulanty, konkrétně SILA-BAC® Kombi, SILA-BAC® Mais Kombi a SILA-BAC® CCM Kombi. Tyto inokulanty obsahují dva bakteriální kmeny Lactobacillus buchneri, které v kombinaci s kmeny Lactobacillus plantarum efektivně zlepšují a urychlují kvasný proces. Tím se výrazně zkracuje doba potřebná ke stabilizaci výsledné siláže, snižují se ztráty sušiny během silážování a zabraňuje se růstu kvasinek a plísní.
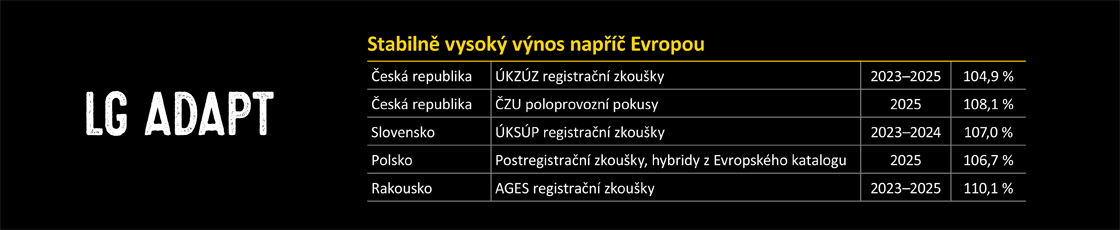
Jak již bylo výše uvedeno, hlavní předností obou inokulantů je velmi rychlé dosažení aerobní stability siláže. To bylo prokázáno opakovanými laboratorními testy, ve kterých byla porovnávána stabilita kontrolní, spontánně fermentované, neošetřené kukuřičné siláže a siláže konzervované inokulantem SILA-BAC® Mais Kombi. Siláže pro tyto testy byly připravovány v laboratorních mini silech. Po deseti dnech fermentace z nich byly odebrány vzorky, které byly umístěny do otevřených termoboxů, a tím vystaveny působení vzdušného kyslíku. Do středu siláží byla umístěna čidla teploměrů a po dobu pěti dní byly monitorovány změny teploty vzorků. Výsledky testů (viz graf) jasně ukazují, že již po deseti dnech fermentace je kukuřičná siláž inokulovaná přípravkem SILA-BAC® Mais Kombi schopna udržet svou teplotní stabilitu minimálně pět dní téměř beze změn oproti kontrolní siláži. Neošetřená kukuřičná siláž se již po 36 hodinách výrazně zahřívá a následně dochází k úplné degradaci silážní hmoty.
Aktivita kvasinek v siláži výrazně zvyšuje ztráty sušiny (živin), způsobuje zahřívání hmoty, snižuje chutnost a příjem krmiva a v neposlední řadě vytváří podmínky pro růst a množení dalších nežádoucích mikroorganismů (plísně, houby hnilobné bakterie). Za přítomnosti kyslíku mají některé druhy kvasinek schopnost metabolizovat kyselinu mléčnou, tím zvyšovat pH siláže a zároveň produkovat aromatické sloučeniny, jakými jsou například estery, etylacetáty a aldehydy, které snižují kvalitu krmiva. Negativní působení kvasinek je možno eliminovat respektováním zásad silážování, dodržením optimální sušiny, technikou a intenzitou odběru siláže a použitím silážních aditiv s osvědčenými kmeny bakterií Lactobacillus buchneri. Siláže ošetřené inokulanty řady Rapid React™ je možno zkrmovat již po deseti dnech fermentace.*
Ing. Vladimír Černík
Pioneer Hi-Bred

Podmínkou efektivní aplikace silážních aditiv je kvalitní aplikátor, v tomto případě aplikátor APPLI-PRO® C500 instalovaný na samojízdné řezačce JD 9600i (Foto archiv firmy)

Siláž je nutné hermeticky uzavřít (Foto archiv firmy)

Kvalita práce na jámě zásadně ovlivňuje výsledek silážování (Foto archiv firmy)












